Существует множество гипотез касательно механизмов, провоцирующих тиннитус, впрочем, ни одна из них до сих пор не была доказана.
Теория диссонирующей дисфункции утверждает, что сигнал тиннитуса появляется во внутреннем ухе при условии, что наружные волосковые клетки повреждены сильнее, чем внутренние.
Теория диссонирующей дисфункции утверждает, что сигнал тиннитуса появляется во внутреннем ухе, когда наружные волосковые клетки повреждены больше, чем внутренние в одной и той же области базилярной мембраны улитки. Повреждения могут быть вызваны чрезмерным шумом, вирусными инфекциями, воздействием некоторых лекарств или просто являться следствием нормального процесса старения (рис. 5).
Внутренние волосковые клетки — рецепторные клетки, ответственные за трансдукцию звука. Они преобразуют механические вибрации структур внутреннего уха (результат проведения звуковых волн из наружного уха в улитку) в электрические импульсы, распространяющиеся по слуховым нервам, после чего происходит восприятие звука.
Большая часть волокон слухового нерва (95%) соединена со внутренними волосковыми клетками и передают электрические импульсы в мозг. Наружные волосковые клетки работают как механические усилители улитки, повышая вероятность улавливания слабых звуков и обеспечивая до 50 дБ усиления. Такое усиление достигается за счет физических вибраций наружных волосковых клеток и может быть оценено методом отоакустической эмиссии. По мере роста громкости внешнего звука, коэффициент усиления постепенно уменьшается.
Когда нейроны в дорсальных кохлеарных ядрах получают возбуждение от внутренних волосковых клеток, а не от поврежденных наружных, то на этом уровне слуховой системы возникает дисбаланс. Это, в свою очередь, вызывает аномальную активность в виде всплесков нейронных разрядов высокой частоты, которые, после усиления в слуховой системе, воспринимаются как шум в ушах.
Если улитка каким-либо образом повреждена, наружные волосковые клетки страдают в первую очередь, затем повреждаются ближайшие внутренние (Chen & Fechter, 2003). Сигналы от обоих типов волосковых клеток сходятся к одной и той же группе нейронов в дорсальном кохлеарном ядре.
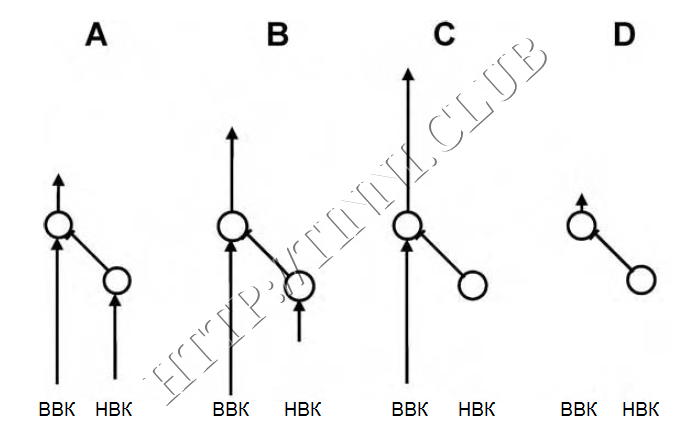
Рисунок 5 – Теория диссонирующей дисфункции.
ВВК – внутренние волосковые клетки, НВК – наружные волосковые клетки. Изображены следующие ситуации. (А) здоровые ВВК и НВК; (B) частично поврежденные НВК и здоровые ВВК; (C) полностью уничтоженные НВК и здоровые ВВК; (D) полностью уничтоженные системы ВВК и НВК. Стрелки изображают сигналы от ВВК и НВК, их длина пропорциональна силе нейронной активности клеток дорсального кохлеарного ядра (ДКЯ) ствола мозга. ![]() — возбуждение,
— возбуждение, ![]() — ингибиция. Интернейрон на пути сигнала от НВК ингибирует клетку на выходе ДКЯ. Обратите внимание на усиление выходного сигнала по мере ужесточения повреждений системы НВК, тогда как при одинаково поврежденных системах наблюдается очень низкий уровень спонтанной активности.
— ингибиция. Интернейрон на пути сигнала от НВК ингибирует клетку на выходе ДКЯ. Обратите внимание на усиление выходного сигнала по мере ужесточения повреждений системы НВК, тогда как при одинаково поврежденных системах наблюдается очень низкий уровень спонтанной активности.
Когда нейроны в дорсальном кохлеарном ядре получают возбуждение от ВВК и не получают от поврежденных НВК, на этом уровне слуховой системы создается дисбаланс. Это, в свою очередь, вызывает аномальную активность в виде всплесков нейронных разрядов высокой частоты, которые, после усиления в слуховой системе, воспринимаются как шум в ушах. Характерная тиннитусу повышенная спонтанная активность была зафиксирована в дорсальном кохлеарном ядре (Kaltenbach & Afman, 2000) и нижнем двухолмии ствола мозга (Chen & Jastreboff, 1995; Jastreboff & Sasaki, 1986; Kwon et al., 1999) (Рис. 6 и 7). Кроме того, в нижнем двухолмии были отмечены всплески спонтанной активности, коррелирующие с тяжестью восприятия тиннитуса (Jastreboff et al., 1999; Kwon et al., 1999).
По меньшей мере два механизма могут выступать в качестве источника нейронной активности, провоцирующей шум в ушах (либо по отдельности, либо оба сразу). Во-первых, амплификация сигнала на любом уровне слуховых путей может привести к усилению естественной колебательной спонтанной активности до уровня, при котором эти колебания начинают восприниматься как звук (тиннитус). Во-вторых, потенциальным источником становятся (даже очень незначительные или точечные) сбои в функционировании или повреждения системы НВК.
По меньшей мере два механизма могут выступать в качестве источника нейронной активности, провоцирующей шум в ушах. Во-первых, амплификация сигнала на любом уровне слуховых путей может привести к усилению естественной колебательной спонтанной активности до уровня, при котором эти колебания начинают восприниматься как звук (тиннитус). Эксперимент со звукоизолированной комнатой (Heller & Bergman, 1953) указал на то, что подобное явление имеет место. Во-вторых, потенциальным источником становятся даже очень незначительные или точечные сбои в функционировании или повреждения системы НВК, которые не всегда могут быть диагностированы такими стандартными слуховыми тестами, как тональная аудиограмма, и о которых сам пациент может не подозревать.
Дисбаланс активности нейронов по-разному воздействует на тип I и тип II волокон слухового нерва, что приводит к всплеску активности на уровне дорсального кохлеарного ядра. После дальнейшего усиления в слуховых путях такая активность может восприниматься как тиннитус.
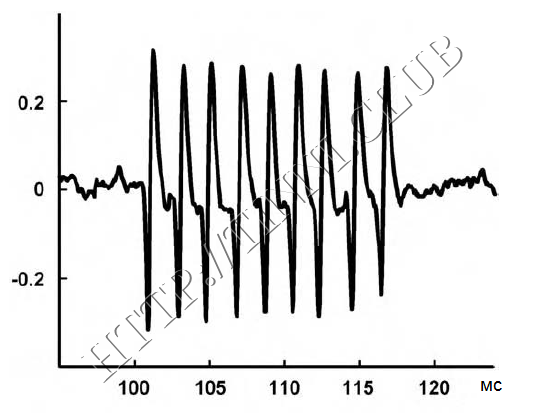
Рисунок 6 — Всплески нейронной активности в нижнем двухолмии у крыс.
Приведена запись спонтанной активности одного нейрона у крысы с тиннитусом, вызванным салицилатами. Частота очень высока, подобная длительная активность обычно наблюдается только при эпилепсии. Вероятнее всего, именно такая активность является причиной тиннитуса (Chen & Jastreboff, 1995).
Возникающий в активности нейронов дисбаланс между I и II типом волокон слухового нерва провоцирует всплеск активности на уровне дорсального кохлеарного ядра, который, после дальнейшего усиления в слуховых путях, может быть воспринят как тиннитус. Из-за тонотопической организации улитки и слуховых путей, локальные нарушения могут привести к восприятию звуков, высота которых соответствует определенной области базилярной мембраны. Когда воздействие прикладывается сразу к нескольким областям базилярной мембраны, результирующий воспринимаемый сигнал может иметь очень сложную природу и представлять возмущения от множества отдельных источников.
Вероятно, комбинация этих двух факторов создает ощущение шума в ушах. Эксперименты на животных показывают, что временная или постоянная потеря слуха (включая дисфункцию НВК) приводит к ощутимому повышению чувствительности у высокого процента нейронов слуховых путей (Gerken, 1979; Gerken, Saunders & Paul, 1984). Снижение количества или повреждение входов в слуховую систему компенсируется на уровне нейронных путей. Последние отчеты подтверждают предположение о том, что степень повреждения НВК имеет прямое отношение к тиннитусу (Kaltenbach et al., 2001; Mitchell & Creedon, 1995).
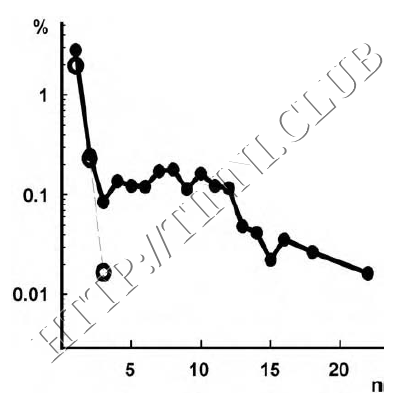
Рисунок 7 – Превалирующая активность у подконтрольных животных и крыс с салицилат-индуцированным тиннитусом.
Вертикальная ось показывает отношение числа последовательностей импульсов установленной длины к общему количеству интервалов, на которых записывался отклик групп клеток на тона соответствующих тиннитусу частот. Горизонтальная ось показывает количество интервалов на одну последовательность импульсов. Обратите внимание, что у подконтрольных животных (незакрашенные круги и пунктирная линия) присутствуют только короткие всплески (не длиннее четырех интервалов) и вероятность их появления низка. У животных с салицилат-индуцированным тиннитусом (закрашенные круги и сплошная линия) можно наблюдать всплески длиной до 23 интервалов, возникающие довольно часто. Данные подтверждают постулат о том, что механизм дезингибиции вызывает нейронную активность, приводящую к тиннитусу. (Chen & Jastreboff, 1995).
Гипотеза диссонирующей дисфункции позволяет объяснить ряд наблюдений. Например, почему воспринимаемая частота тиннитуса, как правило, расположена на аудиограмме в нижней части кривой потери слуха. Эта позиция соответствует области улитки, в которой наиболее остро ощущается разница между поврежденными НВК и нормально функционирующими ВВК.
Приблизительно 20% пациентов с тиннитусом имеют нормальный слух. Это объясняется тем, что изменения слишком незначительны, чтобы быть обнаруженными при помощи стандартной аудиограммы. Тем не менее, они могут привести к диссонансу и запустить компенсаторную реакцию слуховой системы, в результате чего возникает тиннитус.
Почему тогда в 20% случаев тиннитус возникает у людей с нормальным слухом (Davis & El Refaie, 2000)? У пациентов этой группы могут быть микроизменения в системе НВК, которые невозможно диагностировать при помощи обыкновенной аудиограммы. Эти изменения возможно обнаружить при помощи таких тестов, как отоакустическая эмиссия на частоте продукта искажения (ЭЧПИ), объективно оценивающее функционирование системы НВК. Даже небольшие изменения могут привести к диссонирующей активности, вызывая компенсаторную реакцию слуховой системы и провоцируя тиннитус.
У 27% абсолютно глухих людей тиннитус отсутствует, так как при полностью разрушенных как НВК, так и ВВК результирующий дисбаланс ниже, чем когда ВВК продолжают функционировать.
Почему у 27% абсолютно глухих людей тиннитус не наблюдается (Hazell, McKinney & Aleksy, 1995)? Если обе системы (ВВК и НВК) полностью разрушены, результирующий дисбаланс ниже, чем когда одна система продолжает работать, а другая не передает никакого сигнала.
У большинства пациентов тиннитус является побочным эффектом нормальной компенсаторной активности слуховой системы.
У большинства пациентов тиннитус является не более чем побочным эффектом нормальной компенсаторной активности слуховой системы. Система постоянно пытается восстановить гомеостаз и обеспечить наилучший слух, какой только возможно.
В момент прохождения рассогласованного сигнала от улитки к мозгу, как при наличии вышеописанной слуховой дисфункции, так и при ее отсутствии, слуховые пути всегда пытаются компенсировать ущербность входных клеток и скорректировать гетерогенность сигналов, соответствующих соседним частотам. Такой механизм действительно позволяет улучшить слух, но, в то же время, он провоцирует и усиливает нейронную активность, порождающую тиннитус.
Содержание:
28.02.2017
Есть желание поддержать проект? Ознакомьтесь, что вы можете сделать.